
Blog
Medvěd krátkočelý
10.11.2012 22:00
Medvěd krátkočelý (Arctodus simus), je dnes již pravděpodobně vyhynulým druhem pleistocénního medvěda s neobvyklým krátkým čumákem. Medvěd žil asi 3 mil. - 10 tis. let př. n. l. Jednalo se o největšího známého medvěda.
Obsah[skrýt] |
[editovat]Popis

Tento medvěd byl velmi neobvyklé stavby těla. Měl vysoké, lehké nohy, podlerekonstrukčních paleontologů se pohyboval podobným stylem jako velbloud. Měl krátký ocas a malou kulatou hlavu. Tlapy byly široké asi 35 cm, byly tedy přibližně stejně široké, jak měl dlouhý čumák. Byl vysoký asi 1,6 a dlouhý 3,7 metrů, vážil okolo 1100 kg.
Krátký čumák sloužil pravděpodobnému mrchožroutovi velice dobře, má dobrý efekt v lámání morkových kostí. Kdyby kousal kost koncem tlamy, neměl by takovou sílu, jako když umístí kost v tlamě blíž k pantům (páka je tu mnohem větší). Hlavně, když přišel k nějaké mršině pozdě, stále ještě měl z čeho načerpat další energii.
[editovat]Život
Podle výzkumů se spíše než o nebezpečného dravce jednalo o mrchožrouta. Svou velikostí mohl zastrašit jiné potravní konkurenty a zmocnit se jejich kořisti. Lehké, dlouhé nohy v kombinaci s chůzí podobné velbloudí umožňovaly přesun na velké vzdálenosti. Naopak pronásledovat běžící a kličkující kořisti mohl tento medvěd jen obtížně. Dlouhé končetiny neumožňovaly změnit prudce směr běhu.
[editovat]Rozšíření
Tento medvěd se prokazatelně vyskytoval pouze v Americe. Jeho přechod do Asie přes zamrzlou Beringovu úžinu nelze vyloučit, nebyl však doposud potvrzen.
[editovat]Opravdu nežije?
Někteří kryptozoologové se však podle zpráv přicházejících z Kamčatského poloostrova (Rusko), domnívají, že tento medvěd stále přežívá v drsné kamčatské krajině. Pravděpodobnost, že by tomu tak bylo, je však téměř nulová.
Therizinosaurus
10.11.2012 20:19
 Therizinosaurus („Ještěr s kosou“) byl největším dnes známým terizinosauridem. Dorůstal délky 10-12 metrů a hmotnosti 3-6 tun (čímž patřil dokonce k největším známým teropodům vůbec). Tento poměrně podivný dinosaurus s velkými drápy (až přes 1 metr) a pažemi dlouhými až 2,5 metru žil asi před 72-68 miliony let (v období svrchní křídy) na území dnešního Mongolska. Je pravděpodobné, že i Therizinosaurus byl opeřený, stejně jako jeho blízký příbuzný Beipiaosaurus inexpectus.
Therizinosaurus („Ještěr s kosou“) byl největším dnes známým terizinosauridem. Dorůstal délky 10-12 metrů a hmotnosti 3-6 tun (čímž patřil dokonce k největším známým teropodům vůbec). Tento poměrně podivný dinosaurus s velkými drápy (až přes 1 metr) a pažemi dlouhými až 2,5 metru žil asi před 72-68 miliony let (v období svrchní křídy) na území dnešního Mongolska. Je pravděpodobné, že i Therizinosaurus byl opeřený, stejně jako jeho blízký příbuzný Beipiaosaurus inexpectus.
[editovat]Drápy
Jeho drápy mohli být i přes 1 metr dlouhé! Pravděpodobně je používal na obranu před svými nepřáteli.
Moeritherium
10.11.2012 20:11
Moeritheriidae je vymřelá čeleď chobotnatců. Obsahuje pět zástupců jediného rodu Moeritherium. Žili v eocénu. Byli menší než současní sloni, na délku měřili kolem tří metrů, na výšku asi 70 cm. Byli podobní tapírům, svým způsobem života a rostlinnou stravou ale přípomínají spíše hrochy. Zástupci čeledi nejsou přímými předky slonů, jejich nejbližším příbuzným je snad rod Numidotheriumnebo čeleď Deinotheriidae a jí příbuzné rody.
mamenchisaurus
10.11.2012 18:33
Mamenchisaurus („ještěr od potoka Mamen“) byl 27 tun těžký a až 25 metrů dlouhý sauropodní dinosaurus, který žil před asi 160 až 145 miliony let. Jeho fosílie byly nalezeny v Číně.
Zajímavostí tohoto druhu je jeho krk, který je nejdelší mezi všemi známými dinosaury (až kolem 13 metrů). Tvořilo jej 19 obratlů a přes svou délku byl relativně lehký. Nejdelší objevené "krční žebro" tohoto dinosaura (druh M. sinocanadorum) měřilo na délku celých 4,1 metru, takže jde možná o nejdelší známou obratlovčí kost vůbec (nikoliv však nejobjemnější nebo nejtěžší).
Spinosaurus
10.11.2012 18:23
Spinosaurus („Trnitý ještěr“) byl obrovský masožravý dinosaurus (teropod), který žil na území dnešní severní Afriky v křídovýchperiodách albu a cenomanu před asi 95-93 miliony let. V současnosti je považován za největšího masozraveho dinosaura.Podle studie dal Sasso et al., 2006 byl právě tento druh největším známým teropodním dinosaurem, větším než dosavadní držitel primátu Giganotosaurus. Největší udávané rozměry tohoto dravého obra činí 15 (nebo 16-18) metrů délky a kolem 9 tun hmotnosti. Některé odhady posouvají hmotnost až k neuvěřitelným hodnotám 12-19 tun, což je na samotné hranici únosnosti pro dvounohého živočicha. Jenom lebka jednoho exempláře by prý v kompletním stavu byla dlouhá až 2,4 metru (i když oficiální odhady spíš hovoří o délce 1,75 m). Tím by Spinosaurus představoval zdaleka největšího dravého tvora, který kdy chodil po souši (a jehož fosílie již známe).Rekordní velikost
[editovat]Paleobiolpp
Charakteristické jsou pro tento druh dlouhé zádové výběžky obratlů, tvořící jakési „trny“ o délce až přes 2 metry, za života zvířete zřejmě sloužící v podobě kožní plachty, napnuté mezi těmito výběžky, k termoregulaci. Spinosaurus žil nejspíš na březích křídových jezer a potoků a lovil velké sladkovodní ryby, možná se živil částečně i mršinami. Pro to svědčí i utváření jeho zubů a čelistí, vhodných k lovení ryb ve vodě. Obojživelný způsob života potvrzuje také výzkum z roku 2010[1]. Nelze samozřejmě vyloučit ani potravu v podobě živých a snad i mrtvých velkých býložravých dinosaurů, žijících v této oblasti před 95 miliony let.
Čtvrtihory
08.11.2012 21:24

Poslední údobí ve vývoji naší planety označujeme jako kenozoikum neboli novověk Země. Tato éra trvala asi 65 miliónů let a je pro nás zvláště důležitá. Během ní se vyvinula skupina primátů (nehetnatců), z níž jako jeden z posledních článků vznikl i člověk. Kenozoikum tvoří jediné přirozené období, charakterizované vyvrcholením a dozněním mohutného alpínského vrásnění, při kterém vznikla mladá pásemná pohoří, a postupným utvářením a modelováním zemského povrchu až v jeho nynější podobu. Rovněž vývoj života v celém tomto údobí má celkově shodné rysy a obdobný charakter. Geologové dělí kenozoikum na dvě nestejně dlouhá údobí, a to je terciér a kvartér neboli třetihory a čtvrtohory. Třetihory, které zabírají většinu kenozoika, se dále dělí na údobí starší zvané paleogén (39 mil. let) a mladší zvané neogén (24 mil. let). Paleogén členíme ještě na kratší epochy, a to na paleocén, eocén a oligocén. Neogén dělíme pak na miocén a pliocén. Čtvrtohory představují v geologickém čase jen krátký okamžik, asi 2-3 mil. let. Dělíme je na starší - pleistocén a mladší - holocén. Jako samostatný útvar se čtvrtohory oddělují proto, že tvoří údobí geologicky dobře vymezené a svým způsobem, i přes malé geologické stáří, výjimečné.

Posledním a nejkratším geologickým údobím jsou čtvrtohory. Představují sotva začínající epochu, která zatím trvá dva až tři (?) milióny let. V dějinách Země zaujímají mimořádné postavení, a to ze dvou důvodů: 1. teplota periodicky, ale zároveň poměrně rychle klesala, takže se na povrchu souší značně rozšířily ledovce, 2. vyvinul se moderní člověk, který svou přítomností, kulturou i celou činností toto údobí ovlivňuje. Proto se čtvrtohory označují také jako doba člověka neboli antropozoikum.
Geologové je dělí na dvě údobí, a to na starší, pleistocén a mladší, holocén. Tomuto nejmladšímu období, které zatím trvá jen asi 10 000 let, říkáme často též recent neboli doba současná. Čtvrtohory trvají dodnes a za jejich začátek byla dohodnuta doba, kdy se výrazně ochladilo Středozemní moře. Při jejich dělení se často používá členění běžné v archeologii. Doba pleistocenní se označuje jako starší doba kamenná neboli paleolit a doba holocenní se dělí na střední dobu kamennou (mezolit) a mladší dobu kamennou (neolit).
Ve čtvrtohorách nastalo nápadné ochlazení, které bylo vyvrcholením postupného kolísání klimatu, pozorovaného již v pozdních třetihorách. Silné ochlazení na konci třetihor mělo hluboký vliv na existenci a vývoj všeho živého. S klesající teplotou přibývalo atmosférických srážek, které se v chladných ročních obdobích měnily ve sněhové vánice. Vznikaly rozsáhlé sněhové příkrovy, které se postupně měnily v příkrovy ledové až v ledovce. Tak vznikalo zalednění obrovských prostor naší země. Odhaduje se, že v době svého vrcholu, v pleistocénu, pokrýval ledovec 45 mil. km2 souše. V Severní Americe jsou vidět stopy zalednění až ke 40. rovnoběžce. V daleko větším měřítku než dnes byly zaledněny i horské masívy. V té době se také vyhranila velmi výrazná podnebná pásma, a to pásmo arktické, mírné a tropické. Jejich hranice se ovšem podle kolísání průměrné teploty během pleistocénu značně měnily.
Geologové dokázali, že ochlazení ke konci třetihor a na počátku čtvrtohor nebylo trvalé, ale že bylo vystřídáno údobími teplejšími, po nichž nastaly opět doby chladnější atd. Podnebí ve čtvrtohorách se tedy vyznačovalo střídáním chladnějších dob ledových, tzv. glaciálů, a teplejších dob meziledových, tzv. interglaciálů. Během čtvrtohor se ledové doby i jejich interglaciály rytmicky opakovaly v několika velkých cyklech. Bylo zjištěno, že např. v Alpách byly během čtvrtohor nejméně 4 velké doby ledové a 3 údobí meziledová. Rytmické střídání chladnějších a teplejších období probíhalo na obou polokoulích současně.
Během ledových dob se v předpolí severského ledovce rozkládala v šíři 200-300 km tři vegetační pásma, která označujeme jako tundra, step a tajga. V tundře, která se přimykala těsně k čelu ledovce, rostly různé mechy a lišejníky (hlavně lišejník sobí), dále zakrslá polární vrba (Salix polaris), zakrslá bříza (Betula nana), a hlavně dryádka osmiplátečná (Dryas octopetala), podle níž se tundrová flóra ledových dob označuje jako flóra dryasová. Její součástí byl i nízký pěnišník (Azalea procumbens), různé druhy lomikamenů, rožců a jiných. Ve stepním pásmu rostly převážně různé druhy vrb, bříza bílá (Betula alba), místy zakrslé formy borovic (Pinus montana a Pinus silvestris) a vedle nich pak četné typy kvetoucích rostlin. V pásmu tajgy, které bylo zatlačeno poměrně k jihu, rostly různé jehličnany a listnáče.
Měkkýši mají ve čtvrtohorních usazeninách prvořadý význam pro členění mořských i kontinentálních uloženin.
Pro pleistocenní faunu byli charakteristíčtí především savci, a to hlavně chobotnatci (Proboscidea). V nejstarším pleistocénu žil Archidiskodon planifrons a v jižní Evropě Archidiskodon meridionalis. Největší z vyhynulých evropských pleistocenních slonů byl rod Palaeoloxodon, který dosahoval v lopatkách až pětimetrové výšky. Žil v lesnatých porostech během meziledových dob. Ze všech vyhynulých pleistocenních zvířat je nejlépe známý mamut (Mammuthus primigenius), význačný představitel glaciální fauny. Jeho hlavní rozvoj proběhl ve svrchním pleistocénu; žil v tundře.
Současně se slony žili v pleistocénu i nosorožci. Ve starším pleistocénu to byly druhy Dicerorhinus etruscus a Dicerorhinus kirchbergensis, typičtí představitelé interglaciální doby.
K velkým významným pleistocenním savcům patří z lichokopytníků rod Equus - kůň. Tento rod, odvozující se přímo od pleistocenního rodu Pliohippus, vznikl v Severní Americe, odkud se rozšířil přes celou Asii až do Evropy. V Severní Americe však vyhynul už koncem pleistocénu a zavedl ho sem znovu až po objevení Ameriky bílý člověk.
Ze sudokopytníků se v teplých dobách starých čtvrtohor vyskytoval v Evropě i hroch (Hippopotamus); jeho zbytky byly nalezeny až v Anglii. Sudokopytníci byli ve starších čtvrtohorách zastoupeni také rodem Cervus (jelen). Velmi pozoruhodným představitelem tohoto rodu byl obrovský jelen irský Megaceros giganteus, jehož parohy podobné parohům daňka měly v rozpětí až 3 m.
V pleistocénu žily v Evropě hojně šelmy, především medvědi. V předposlední době ledové u nás žil obrovský medvěd jeskynní (Ursus spelaeus); vymřel během poslední doby ledové. Ze staršího pleistocénu je znám v Evropě šavlozubý tygr (Machairodus), který měl nápadně velké špičáky. Ve středním a svrchním pleistocénu žil v jižní i střední Evropě jeskynní lev Panthera spelaea, který měl téměř stejnou kostru jako dnešní lev, ale vzhledem (exteriérem) se od něho velmi lišil. Z dalších šelem se u nás vyskytovali během ledových dob hyena, liška, vlk, stepní tchoř, rosomák aj. Během pleistocénu u nás byli velmi hojní i hlodavci (Rodentia). Z této doby jsou také známí lumíci (rod Myodes), syslové (Citellus), bobr (Castor fiber) aj. Se zbytky savců se vyskytují v uloženinách ledových dob také zbytky ptáků, především kura rousného, kura alpského, některých sov, pěnkav aj.
Během čtvrtohor vyvrcholil vývojový proces, který začal u primitivních primátů v paleocénu a skončil vznikem moderního člověka v současnosti. Člověk vzešel z okruhu úzkonosých opic Starého světa. Někdy koncem starých třetihor se od nich odštěpila nová vývojová linie, nadčeleď Hominoidea, do které patří jak lidoopi, tak lidé. Tato nadčeleď se dělí do čeledí: Oreopithecidae, Pliopithecidae, Pongidae a Hominidae (hominidi).
Čeleď Oreopithecidae je vymřelá. Jediným představitelem je Oreupithecus bamboli, který dosahoval velikosti šimpanze.
Čeleď Pliopithecidae zahrnuje řadu nálezů z Afriky, Evropy a Asie. Z fosilních rodů je nejvýznamnější Pliopithecus. Pliopithecus dosahoval velikosti dnešního gibbona.
Čeleď Pongidae má mnoho rodů, které řadíme do tří podčeledí označených podle typických rodů: Dryopithecinae, Ponginae a Gigantopithecinae. Do podčeledi Dryopithecinae zahrnujeme dnes rody Dryopithecus, Aegyptopithecus a Propliopithecus, které vznikly pravděpodobně v oligocénu. Čelisti, zuby a některé dlouhé kosti ukazují na opici velkou asi jako šimpanz, ale menší, slabší postavy.
K počeledi Ponginae patří žijící orangutan, šimpanz a gorila. Předky afrických příslušníků této podčeledi neznáme.
K podčeledi Gigantopithecinae patří vymřelý, velmi nápadný a zajímavý rod Gigantopithecus, jehož zbytky byly objeveny v pleistocénu Číny a v Indii; byl větší než gorila.
Nejvýše organizovanou a nejdůležitější čeledí primátů je čeleď Hominidae (lidé). Patří k ní i člověk a jeho předchůdci. K rozvoji této čeledi stačilo asi 14 miliónů let; k rozvoji rodu Homo bylo zapotřebí pouze 2-3 miliónů let. V současné době řadíme do této čeledi (C. P. Groves, 1970) čtyři rody, a to Ramapithecus, Australopilhecus, Paranthropus a Homo.
Hlavní dokladový materiál rodu Ramapithecus pochází z Indie a Číny, odkud byl popsán Ramapithecus punjabicus. Ramapithecus byl menší než dnešní člověk (asi 110 cm vysoký) a pohyboval se po dvou zadních končetinách. Je to zatím nejstarší známý předchůdce člověka; žil asi před 12 milióny let ve stepích a lesostepích.
Rod Australopithecus dnes slučuje četné nálezy z východní a jižní Afriky. Australopitekové byli asi 120 cm vysocí, pohybovali se po dvou končetinách, měli vzpřímenou postavu a mozkovnu s maximální kapacitou kolem 454 ccm (max. kapacita mozkovny lidoopů je 450 ccm). Žili v rovinách a stepích a živili se převážně masitou potravou a plody. Pravděpodobně již lovili i větší zvířata, jako koně, žirafy aj. Na lov se vydávali v tlupách, v nichž se uplatňovala určitá spolupráce. Celkově lze říci, že australopitekové představují další fázi ve vývoji člověka a že byli dobře vyhraněnou skupinou, vývojově mnohem pokročilejší než skupiny předchozí.
Podobně jako u rodu Australopithecus spojuje i rod Paranthropus několik dříve různě jmenovaných nálezů. V současné době se v tomto rodu rozlišují dva druhy: Paranthropus robustus (Broom, 1938) a Paranthropus boisei (Leakey, 1959); oba pocházejí z Afriky. Paranthropus byl ve srovnání s australopitekem vyšší a celkově mohutnější. Žil v lesnatém terénu a živil se převážně rostlinnou potravou. Lebku měl masívnější a těžší. Dnes převládá názor, že tento rod byl specializovanou býložravou vývojovou větví čeledi Hominidae, která žila ve stejné době jako Australopithecus a Homo habilis a vyhynula někdy na počátku středního pleistocénu. Podle skutečného datování žili zástupci rodu Paranthropus v rozmezí 2,7-1,5 miliónu let.
V rodu Homo jsou zahrnuty všechny fosilní i žijící populace člověka. Patří sem Homo habilis, Homo erectus (syn. Pithecanthropus erectus) s dalšími poddruhy.
Nejstarší známý představitel rodu Homo je druh Horno habilis. Původně byl označován jako "pokročilý" Australopithecus. Tento první pravý člověk dosahoval výšky kolem 130 cm a váhy asi 30-42 kg. Lebka byla zaoblenější než u australopiteka, čelo ploché, nadočnicové valy - hlavně u mužů - mohutné; bradový výběžek nebyl vytvořen. Průměrná kapacita mozkovny činila 641 ccm, a byla tedy větší než u australopiteka (454 ccm). Homo habilis žil asi před 1 750 000 lety. Měl vzpřímenou postavu a byl dobře přizpůsoben k pohybu po dvou končetinách. Vyráběl a používal kamenné nástroje.
Mozkovna Homo erectus je na rozdíl od australopiteka mírně klenutá, kapacita lebeční se pohybuje v rozmezí 750-1250 ccm. Čelisti jsou silné, obličej mohutný. Podle tvaru nalezené stehenní kosti byla jeho postava vzpřímená a dosahovala výšky 165-170 cm. Hlava byla předkloněná, obličej byl hrubě modelovaný s malým širokým nosem a se silnými vystouplými čelistmi. Čelo bylo nízké, nazad ubíhající, nadočnicové oblouky byly nadměrně vyvinuty, takže tvořily nad hluboko vsazenýma očima jakousi stříšku. Spodní čelist byla bez brady.
Až donedávna se považoval za předchůdce člověka neandertálec. První zbytky byly objeveny v r. 1856 ve Feldhofské jeskyni v Neanderově údolí řeky Düsselu, východně od Düsseldorfu v NSR. Původně byly popsány jako Homo neanderthalensis King, 1864. Dnes se neandertálec považuje jen za vymřelý poddruh druhu Homo sapiens; správný název je tedy Homo sapiens neanderthalensis.
Vznikem druhu Homo sapiens vyvrcholil vývoj řádu Primates, který trval z paleontologického hlediska poměrně velmi krátkou dobu, asi 65 miliónů let.
Druhohory
08.11.2012 21:23
 Giganotosauři - větší než tyrannosaurus
Giganotosauři - větší než tyrannosaurus Spinosaurus - větší než tyrannosaurus a větší než giganotosaurus, 15 m
Spinosaurus - větší než tyrannosaurus a větší než giganotosaurus, 15 m Suchomimus
Suchomimus Baryonyx
Baryonyx

 Ornithomimus
Ornithomimus
 Therizinosaurus
Therizinosaurus Oviraptor
Oviraptor Tyrannosaurus - NEbyl největším suchozemským predátorem
Tyrannosaurus - NEbyl největším suchozemským predátorem Velociraptoři
Velociraptoři Argentinosaurus - délka: 40-45 m
Argentinosaurus - délka: 40-45 m Triceratops
Triceratops
 Ankylosaurus
Ankylosaurus Iguanodon
Iguanodon
 Parasaurolophus
Parasaurolophus Pachycephalosaurus
Pachycephalosaurus Deinosuchus - 15 m
Deinosuchus - 15 m
 Mosasaurus - 18 m
Mosasaurus - 18 m Kronosaurus
Kronosaurus Tylosaurus
Tylosaurus Elasmosaurus - 14 m
Elasmosaurus - 14 m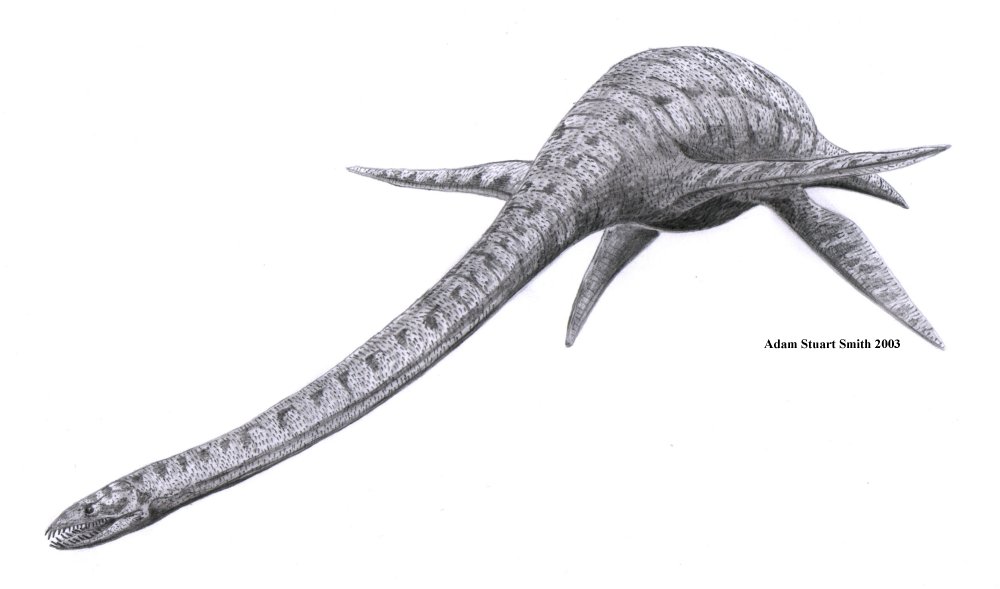

 Pteranodon
Pteranodon
 Quetzalcoatlus - délka 7,5m, rozpětí křídel - 14 m
Quetzalcoatlus - délka 7,5m, rozpětí křídel - 14 m
Třetihory
08.11.2012 21:19
 Gastornis
Gastornis

 Didolodus
Didolodus Phenacodus
Phenacodus Mesonyx
MesonyxDruhohory
08.11.2012 21:18
(230 - 63 mil.let)
trias, jura, křída
trias (230 -181 mil.let)
živočichové - vývoj krytolebců, rozvoj plazů (dinosaurů) - vyvrcholení v juře, v moři měkkýši (hlavonožci - amoniti)
rostliny - přesličky a kapradiny nových rozměrů, hojnost nahosemenných rostlin - cykasovité, jehličnaté
jura(181 - 135 mil. let)
- této době se začal velký kontinent rozpadat
- rozšíření mělkých moří na kontinentech - docházelo zde k usazování velkého množství materiálu (písky -› pískovec, uhličitanové horniny -› křída)
- začalo Alpinsko - himalajské vrásnění
živočichové - velký rozvoj hlavonožců (amonitů), plazi - býložravci (Stegosaurus), masožravci (Allosaurus), první ptáci (Archaeopteryx), primitivní savci
rostliny - jehličnany, cykasy, drobné kapradiny a přesličky
křída (135 - 63 mil. let)
živočichové - postupně vymřely velcí plazi (býložravec - Iguanodon, masožravec - Tyranosaurus), rozvoj hadů, ptáků, vačnatců, hmyzožravců
rostliny - krytosemenné rostliny .
Prvohory
08.11.2012 21:00
Prvohory trvaly celkem 345 miliónů let. Geologové dělí tuto éru na šest period. Kambrium, ordovik, silur a devon (starší prvohory) a karbon a perm (mladší prvohory). Prvohory v podstatě tvoří dva velké vývojové cykly. První (kaledonský) skončil v polovině devonu; druhý (variský nebo hercynský) skončil koncem permu.
Někteří geologové se domnívají, že koncem prekambria existoval jediný obrovský kontinent zvaný Pangea, který byl úplně obklopen praoceánem, ale který se brzy rozpadl na několik částí. Počátek prvohor charakterizuje velká záplava, při níž moře vniklo do rozsáhlých oblastí, jež byly koncem prekambria soušemi. V prvohorách se jednotlivé, horotvornými pochody poněkud zvětšené části původního celistvého prakontinentu nejprve vzájemně vzdalovaly, ale brzy se opět pozvolna sunuly k sobě tak, že koncem paleozoika byla Pangea opět zcelená. Od počátku druhohor docházelo postupně k novému rozčleňování Pangey na jednotlivé kontinenty sunoucí se od sebe tak, že posléze zaujaly svou dnešní polohu. Mezery mezi nimi byly zality oceánem. Tuto domněnku o stěhování kontinentů poprvé jasně definoval r. 1912 německý profesor Alfred L. Wegener. Jeho teorie byla později zpřesněna a doplněna mnohými novými poznatky. Dnes ve své nové formě je označována jako domněnka o pohybu desek zemské kůry a je téměř všeobecně uznávána. Pangea se v první fázi rozdělila na superkontinenty. Na severní polokouli to byla Laurasie a na jižní polokouli Gondwana. Mezera mezi nimi byla zalita oceánem, který byl přechůdce pozdějšího velkého středozemního moře zvaného Tethys.
Následkem kaledonského vrásnění se v dalším vývoji během siluru připojily k severnímu kontinentu rozsáhlé oblasti, které dříve zaplavovalo moře. Tak se v devonu pozvolna zvedal a formoval rozsáhlý severní kontinent, jehož velmi nepravidelný a členitý povrch vlivem horkého a suchého podnebí silně větral. Horské masívy se rozpadaly a uvolňováním sypkého materiálu vznikaly mohutné vrstvy usazenin. Rozsáhlá devonská pevnina se podle převládající barvy sedimentů označuje jako Starý červený kontinent. Na něm se vyvinuly četné nové typy suchozemských rostlin a v Grónsku na něm byly zjištěny i zbytky prvních suchozemských obratlovců, primitivních obojživelníků (krytolebců). Starý červený kontinent měl tedy pro vývoj života zvláštní význam. V pozdějších prvohorách došlo na Gondwaně, k níž patřila Jižní Amerika, téměř celá Afrika, Madagaskar, Indie a Antarktida, k rozsáhlému zalednění. Její rozpad začal až počátkem druhohor. Na konci prvohor moře ustoupilo a hercynské horotvórné pochody zvolna ustaly. V té době vymřelo mnoho primitivních typů rostlin a zvířat.
Během prvohor se postupně vyvinuly různé skupiny rostlin. V prvním počátečním období, až do siluru, ještě převládaly mořské řasy. Později, do konce karbonu, se hojně vyvíjely sporonosné výtrusné rostliny (kapradiny). Ke konci prvohor, zvláště v druhé polovině permu, převládaly hlavně rostliny nahosemenné.
Zvířena, která se objevila na počátku prvohor, měla již pevné kostry, schránky nebo štíty, které poskytují dobrou ochranu a oporu choulostivým částem těla a slouží jako zásobárna různých anorganických (neústrojných) látek umožňujících rozměrnější stavbu těla. Živočichové s pevnými schránkami dosáhli v kambriu rychlého rozvoje. Protože možnost zkamenění pevných schránek a koster byla daleko větší než u zvířeny prekambrické, zachovaly se hned z počátku kambria četné druhy bezobratlých živočichů a z konce tohoto útvaru již i první pozůstatky obratlovců (bezčelistní - Agnatha).
Počátek prvohor - kambrium
Nejstarší útvar prvohor kambrium trval 70 miliónů let. Jeho počátek oznamoval pokles pevninských bloků a rozsáhlá mořská záplava (transgrese), která zasáhla hluboko do starých štítů a vrcholila uprostřed kambria. Úroveň moře a rozsah mořské záplavy se často měnily, protože kůra zemská byla zřejmě v pohybu. Záplavy postihovaly hlavně severní polokouli, kdežto na jižní polokouli zůstávala Gondwana téměř vytrvale vynořená. Koncem kambria moře z četných oblastí ustoupilo, ale v Severní Americe dosáhla tehdy mořská záplava vrcholu. Na počátku kambria ještě doznívalo chladnější podnebí, ale brzy nastalo zřetelné oteplení a koncem kambria, kdy moře opět ustoupilo a plocha souší se podstatně zvětšila, převládlo podnebí teplé a spíše suché.
Dobře zachované zbytky rostlinstva jsou v kambriu velmi vzácné. Hojnější jsou pouze zbytky různých řas. Suchozemské rostlinstvo není téměř známo, jen ojedinělým nálezům se někdy připisuje suchozemský původ. Je však pravděpodobné, že již v této době začaly primitivní rostliny zvolna pronikat na souš.
Rostliny, které začaly růst na souši, musely se pozvolna přizpůsobovat novým, daleko složitějším podmínkám. Části rostlinného těla, které rostly na souši, musely mít silnou pokožku, která je chránila před přílišným odpařováním vody. Aby se mohly vyšší rostliny přizpůsobit pobytu na souši, potřebovaly jednak četná nová složitá zařízení ve vlastním těle, jednak dostatečně mocné vrstvy výživné půdy. K jejímu vytvoření však byly nezbytné nejen četné mikroorganismy, baktérie, sinice, řasy a houby, ale i půdní živočichové. Jejich činnost i jejich odumřelá těla měnily usazené horniny pozvolna v úrodnou půdu, která mohla poskytnout obživu prvotním vyšším rostlinám.
Jak již bylo řečeno, ve vrstvách spodního kambria se náhle objevuje velké množství koster či schránek bezobratlých živočichů., které silně kontrastuje s nesmírně vzácnými nálezy fauny prekambrické. Fauna v kambriu je již tak dobře rozlišena, že v ní lze najít zástupce téměř všech kmenů bezobratlých, a dokonce i prvé zástupce obratlovců. Tento vysoce rozvinutý stupeň života v kambriu však nutně předpokládal dlouhý vývoj v prekambriu.
Nejvýznačnější skupinou živočichů v kambriu jsou bezesporu trilobiti, velmi dobře známí laikům a nesmírně oblíbení u sběratelů zkamenělin. Patří do rozsáhlého kmene členovců, k nimž náleží přes 60 % známých živočišných druhů. Jejich primitivní předkové se vyvinuli během pozdního prekambria pravděpodobně z jednoduchých mořských kroužkovitých červů. Již v nejstarším kambriu dosáhli trilobiti překvapivého rozvoje.
Dalšími velmi rozšířenými obyvateli kambrických moří byli archeocyatidi (Archaeocyatha). Měli kornoutovitou kostru a zastávali v mořích roli pozdějších korálů. Dnes je klademe do blízkosti vápenitých hub.
Velmi hojně se v kambriu vyskytují i různé skupiny ramenonožců (Brachiopoda). Jejich podíl mezi kambrickou faunou činí asi 30 %. Jejich dvouchlopňové schránky (hřbetní a břišní) připomínají misky mlžů. Ramenonožci tvoří samostatný kmen. Jejich jméno se odvozuje od párovitých ramen pokrytých brvami, jimiž živočich dýchal. Ramena byla umístěna po obou stranách ústního otvoru a sloužila také k přihánění vody s potravou do úst. Druhy v kambriu nejrozšířenější měly schránky většinou z chitinovité hmoty a u mnohých vybíhal ze zvláštního otvoru pod vrcholem břišní misky silný, svalnatý stvol, jímž se ramenonožec připoutával ke dnu.
Třetí místo mezi kambrickou faunou patří ostnokožcům (Echinodermata). Jejich zástupci patří k dávno vyhynulým rodům. Měli soustavu vodních trubic, dutin a panožek, v nichž kolovala mořská voda. Schránky některých primitivních forem byly obvykle složeny z četných, nepravidelně uspořádaných destiček a na spodní straně těla byl stonek, jímž se živočich příchycoval k podkladu.
Měkkýši (Mollusca) se v kambriu vyskytují vzácně. Na Dálném východě byli nalezeni primitivní hlavonožci. Zřídka byli objeveni i primitivní mlži. Břichonožci (Gastropoda) jsou zastoupeni jen primitivními, široce mísovitými nebo ještě neúplně stočenými ulitami. První zástupcí hlavonožců (Cephalopoda) byli zjištěni nedávno ve svrchním kambriu východní Asie. V kambrických usazeninách se však hojně vyskytují zástupci vyhynulých hyolitů. Ve svrchnokambrických usazeninách Severní Ameriky byly nalezeny drobné šupiny, bezpochyby nejstarších obratlovců, patřících do skupiny bezčelistných (Agnatha).
Kambrická fauna se šířila na okrajích mořských pánví trvalého charakteru a obydlovala pouze mělké oblasti moří, kde se přizpůsobovala nejrůznějším podmínkám prostředí, např. teplotě, slanosti mořské vody, vlivu mořských proudů aj.
Ordovik
Druhý útvar starších prvohor, ordovik, trval zhruba 55 miliónů let. Jeho počátky charakterizují opět horotvorné pohyby tzv. sardinské fáze kaledonského vrásnění a rozsáhlá mořská transgrese. Konec ordoviku je signalizován ústupem moře a v některých oblastech i výrazným vrásněním. Moře byla v ordoviku značně rozšířena, souš byla spíše plochá, jednotvárná. Mezi stabilními částmi pevninských bloků se často vytvářely hlubší pánve; v některých z nich byla i sopečná činnost.
Podnebí v ordoviku jeví již náznaky podnebných pásem; např. v severní Evropě bylo podnebí teplé a v severní Africe chladnější.
Zbytky flóry (květeny) jsou v ordoviku zatím nesmírně vzácné. Z některých částí světa, např. z Estonské SSR byly popsány zbytky fosilních řas a spor, z Čech pak první cévnaté rostliny Boiophyton pragense a Kreiciella, které se považují za první rostliny suchozemské.
I když hojnější zbytky vyšších rostlin zatím neznáme, je nutné považovat ordovik za údobí, v němž se z valné části jejich vznik připravoval a utvářel. Výsledky tohoto vývoje směřujícího k vyšším rostlinám jsou totiž zřetelně zachovány v siluru, kde již nalézáme dobře zachované zbytky skupiny Psillophyta. Přechodné typy mezi řasami a cévnatými rostlinami však vyhynuly a nedochovaly se.
Vznik vyšších rostlin jistě podporovalo příznivé podnebí a hojně rozšířené, celkem ploché souše.
Právě tak jako v mořích kambrických byli i v mořích ordovických hojně rozšířeni trilobiti, kteří zde dosáhli vrcholu svého rozvoje.
Vedle trilobitů byli v ordoviku velmi hojní ramenonožci. Počátkem tohoto útvaru sice zaniklo mnoho starších typů, ale vznikly i četné tvary nové.
Velmi hojní jsou v ordovických sedimentech graptoliti. Dosáhli v té době celosvětového rozšíření a jsou nesmírně důležití pro porovnání stejně starých vrstev usazenin v různých oblastech a pro jejich zařazování do mezinárodní stratigrafické stupnice. Graptoliti tvoří samostatnou vymřelou třídu polostrunatců (Hemichordata). Jsou to vyhynulí drobní, v kolonii pospolitě žijící mořští živočichové. Byli buď přisedlí, nebo byli přichyceni závěsným vláknem na volně plovoucích řasách. Některé rody se vznášely ve vodě neseny zvláštními měchýřkovitými plováky.
Kostry graptolitů se nejčastěji zachovaly v tmavých břidlicích jako lesklé, pilovitě vrubované nárysy. Graptoliti žili v mořích na celém světě, rychle se přizpůsobovali danému prostředí a vytvářeli značné množství poměrně krátce žijících, zato však hojně rozšířených rodů a druhů. Tytéž rody, a dokonce i druhy lze nalézt ve vrstvách stejného stáří na místech vzdálených od sebe i tisíce kilometrů.
V ordoviku nastal také první větší rozvoj měkkýšů. Mlži již byli zastoupeni všemi základními řády. V plném rozvoji byli hlavonožci loděnkovití, kteří měli většinou dlouhé kuželovité schránky buď přímé, nebo stočené. Některé druhy loděnkovitých dorůstaly až několika metrů délky. V ordoviku už nalézáme také první korálnatce, hojní byli i ostnokožci. Zvláště rozšířeni byli jablovci (Cystoidea), jejichž schránka měla tvar kulovitý, vakovitý i zploštělý a tvořil ji různý počet nepravidelně uspořádaných destiček.
Ordovik je útvarem, v němž nalézáme již hojnější nesporné doklady existence zástupců nesmírně důležitého podkmene obratlovců (Vertebrata). Jsou to izolované, drobné kosterné destičky z rozpadlých krunýřů rybovitých praobratlovců, kteří dosud neměli vyvinuté čelisti. Zahrnujeme je rovněž do skupiny bezčelistnatců (Agnatha).
Polostrunatci (Hemichordata) jsou mořští živočichové, představující jednu z mladších vývojových skupin červů. Utváření jejich těla a některé jiné znaky staví celou tuto skupinu jednak do blízkého příbuzenství s ostnokožci, jednak do těsné blízkosti pravých strunatců (Chordata).
Ať už se tedy první strunatci vyvinuli z kterékoliv skupiny, je téměř jisté, že se jejich prvotní formy značně podobaly dnes žijícímu kopinatci (Amphioxus). Měly tělo po celé délce článkované a to jim umožňovalo větší pohyblivost.
O životním prostředí prvních obratlovců se zatím vedou spory. Část badatelů se domnívá, že žili ve vodách sladkých, jiní naopak tvrdí, že žili v moři. Spor dosud nebyl jednoznačně vyřešen, ale zdá se velmi pravděpodobné, že se první obratlovci dostali z moře do sladkých vod již během kambria. Zřejmě to bylo možné teprve tehdy, když se ve sladkých tekoucích vodách vyskytovaly byť velmi vzácně první rostliny, které jim mohly poskytnout část nutné potravy.
Hojnější zbytky nejprimitivnějších obratlovců nalézáme tedy v ordoviku. Patří do skupiny bezčelistných (Agnatha). Jejich těla dosahující délky kolem 10-40 cm byla zčásti chráněna pevnými kostěnými deskami. Místo úst měli bezčelistní na spodu hlavy zvláštní otvor, jímž nasávali drobnou potravu. Známe dvě skupiny; první, štítohlaví (Cephataspidomorhhi), má dosud žijící zástupce (např. mihule), druhá, štítoploutví (Pterashidomorphi), je zcela vymřelá a právě k ní patří nejstarší známé zbytky obratlovců.
Silur
Třetím útvarem starších prvohor je silur. Byl kratší než ordovik, trval asi 45 miliónů let. Jeho spodní hranici vyznačují v mnoha oblastech výrazné horotvorné pohyby (takonská fáze kaledonského vrásnění). Pouze na počátku bylo mořské dno v četných oblastech, např. v Evropě, vyzdviženo. Celkově však mořská záplava po většinu siluru trvala.
Na konci siluru moře z mnoha oblastí ustoupilo a mořské dno se začalo opět zvedat působením rozsáhlých horotvorných pochodů další (ardenské) fáze kaledonského vrásnění. Toto kaledonské vrásnění bylo nejsilnější v severní Evropě. Podél okrajů baltského štítu se z hlubokých mořských příkopů vyvrásnila nová vysoká pásemná pohoří, která se táhla v mohutném oblouku z Anglie přes Skotsko do Norska a odtud přes Špicberky do severního Grónska. Tato pohoří jsou známa pod názvem Kaledonidy. Se silurskými horotvornými pohyby souvisely i procesy vulkanické, které se např. ve středních Čechách projevily podmořskými výlevy diabasových láv.
Podle výskytu četných korálů, které lze poněkud vzdáleně porovnávat s dnešními korálovými útesy tropických moří, můžeme se domnívat, že v době silurské bylo podnebí poměrně teplé. Pás subtropického podnebí sahal mnohem severněji než dnes. Zhruba se již rýsují dvě podnebná pásma, a to teplé a studené.
Život v siluru byl již plně rozvinutý. Měl své vyhraněné charakteristické znaky a lze říci, že představoval význačnou etapu v celkovém vývoji života na Zemi. Vedle téměř všech skupin bezobratlých se dosti rozšířili i obratlovci. Rovněž flóra se rozmáhala a vedle velkého rozvoje řas byly již hojnější i vyšší rostliny.
Četné mikroskopické a makroskopické zbytky řas dokazují, že řasy v této době byly již značně rozšířené. Hojněji se objevily i nejprimitivnější rostliny cévnaté (Psilophyta). Jejich tělo nebylo často ani rozlišeno ve stonky, listy a kořeny.
Mořské usazeniny silurského útvaru jsou velmi bohaté na rozmanité skupiny bezobratlých živočichů. Zvláště velký rozvoj lze pozorovat u měkkýšů (Mollusca), kteří byli v siluru jednou z hlavních složek mořské zvířeny. Četné rody z této doby připomínají již rody současné.
Vývoj měkkýšů byl dlouho nejasný. K jeho objasnění přispěla zčásti i náhoda.
Třebaže paleontologové nashromáždili množství důkazů o kmenovém vývoji a prastarém původu měkkýšů, nemohli své výzkumy doložit studiem měkkých částí, protože ty se na fosilním materiálu nezachovaly.
V siluru dosáhla vrcholu svého rozvoje i jiná skupina měkkýšů, a to loděnky (Nautiloidea). Většinou se jejich schránka podobala protaženému přímému nebo jen mírně prohnutému kuželi. U některých druhů dosahovala délky až 4,5 m. Během vývoje se schránka různě ohýbala, až posléze vznikly formy se schránkami úplně stočenými. Takové druhy byly zřejmě nejlépe připraveny pro boj o život, a proto vzácně, např. loděnka hlubinná (Nautilus pompilius), přežily až do současné doby. Mnoho druhů loděnkovitých však vyhynulo již v siluru.
Zvláštní pozornost v silurské fauně je třeba věnovat ještě dvěma skupinám. Jsou to již zmínění graptoliti a láčkovci.
Láčkovci (Coelenterata) se v siluru značně - rozšířili. Zvláštního významu nabyli korálnatci (Anthozoa), představovaní jednak skupinou korálů drsnatých (Rugosa), jednak skupinou korálů deskatých (Tabalata). Korálnatci jsou mořští živočichové přisedlí ke dnu, žijící buď osamoceně, nebo v koloniích. Jejich kolonie často dorůstaly značných rozměrů a vytvářely buď větší vápenité kostry, nebo velké kruhovité trsy, které byly pevně přisedlé k podkladu.
Vedle korálnatců byla v siluru hojně rozšířena skupina stromatopor (Stromatoporoidea). Vylučovaly pevné vápenité kostry, čočkovitého, bochníkovitého, válcovitého nebo plátovitého tvaru často až 2 m v průměru. Spolu s kostrami korálnatců představují hlavní stavební kameny silurských vápenců.
Během siluru pokračovali v rychlém rozvoji mnozí členovci (Arthropoda), např. obrovští různorepí (Eurypterida). Byli to význační obyvatelé silurských moří, kteří měli vztahy k dnešním ostrorepům (Xiphosura) a pavoukovitým (Arachnida). Některé druhy dorůstaly délky až 3 m a patřily k největším bezobratlým živočichům své doby. Různorepí měli obvykle silná klepeta, ale poměrně malé končetiny. Obývali mělčí části moře i vody brakické (smíšené sladko-slanovodní).
Místy bylo mořské dno doslova přeplněno pestře zbarvenými lilijicemi (Crinoidea, kmen Echinodermata). Ke dnu byly připevněny dlouhým stonkem, na jehož opačném konci bylo tělo živočicha, podobné lilii.
Silný rozvoj zaznamenali i ramenonožci. Zejména v mělkých mořích patřili k nejhojnějším živočichům.
Zatímco živočišné skupiny, které jsme právě uvedli, pokračovaly rychle ve svém rozvoji, jiné skupiny již ve vývoji zaostávaly, nebo dokonce pozvolna vymíraly (např. některé čeledě trilobitů, ostnokožců aj.).
Silur však nebyl jen obdobím rozvoje bezobratlých. Pocházejí z něho také nálezy celých koster již výše uvedených praobratlovců (Agnatha). Měli chrupavčitou vnitřní kostru a tělo z valné části chráněné pevným vnějším krunýřem z četných kostěných destiček. Čelisti jim chyběly, ale na spodní straně hlavy měli malá kruhovitá nasávací ústa a za nimi řadu žaberních štěrbin. Na svrchní straně hlavy byly oči a lichý nosní otvor. U některých se vyvinuly 1 -2 hřbetní ploutve a nesouměrná ploutev ocasní. Tito primitivní obratlovci byli pro vědu velkou záhadou a trvalo dlouho, než byla objasněna jejich anatomie.
Vedle primitivní skupiny bezčelistných (Agnatha) se koncem siluru objevili i první zástupci obratlovců čelistnatých (Gnathostomata), kteří již měli vyvinutou spodní a svrchní čelist. Byly to ryby trnoploutvé (Acanthodii).
Devon - doba ryb
Posledním útvarem starších prvohor je devon. Byl delší než silur, trval asi 50 miliónů let. Celý tento útvar byl nabitý důležitými událostmi, nesmírně významnými pro vývoj organismů. Bylo to neklidné údobí. V jeho první polovině úplně doznělo kaledonské vrásnění. V té době vznikla na severní polokouli obrovská pevnina. Vlivem teplého a suchého klimatu zvětral její skalní podklad a tyto zvětraliny, nápadně červeně zbarvené oxidem železa, se hromadily v četných pánvích v deltách řek a na rozsáhlých nivách. Charakteristické sedimenty této doby označili angličtí geologové jako staré červené pískovce (Old red sandstone). Podle nich se tato devonská prapevnina nazývá Starý červený kontinent.
V té době trvala ve středních Čechách nepřerušená mořská záplava a probíhal nepřerušený vývoj různých ústrojenců. Právě proto jsou střední Čechy klasickou oblastí, která byla zvolena za světový standard pro sledování a vymezení hranice mezi silurem a devonem.
Vznik nové obrovské pevniny na severní polokouli i vznik nových pásemných pohoří měl za následek také vznik podnebných pásem, jak tomu bylo několikrát již dříve. V obvodu Červeného kontinentu bylo (podle nalezené květeny) podnebí teplé a suché. Koncem devonu se však množství srážek zvýšilo a následkem toho se zrychlil rozvoj suchozemských cévnatých rostlin, takže místy už vznikly první uhelné sloje.
Na rozvoji života v devonu měly rozhodující vliv dvě důležité události; jednak vznik obrovského Starého červeného kontinentu, jednak vznik rozsáhlých mělkých teplých moří s hojnými zálivy, které byly ideálním prostředím pro rozvoj obratlovců, hlavně ryb. Proto se také často označuje devon jako doba ryb. Nová pevnina na severní polokouli s četnými řekami, lagunami, jezery i močály, s rozvíjející se suchozemskou květenou a příznivým klimatem byla bezesporu nejvhodnějším prostředím, kde se posléze vyvinuli první čtvernožci dýchající vzduch. Během dlouhé doby devonské se však také pozoruhodně rozvinulo i rostlinstvo. Na sklonku tohoto údobí se již vytvořily mohutné porosty, první suchozemské pralesy. Vznikly výše organizované kapraďorosty, vytvořily se i první typy stromovité. Poprvé se setkáváme se zajímavým jevem, že rostlinstvo ve svém vývoji poněkud předbíhá vývoj živočichů.
Na mnoha místech Starého červeného kontinentu vznikaly rozsáhlé oblasti úrodné půdy, kde pak došlo k rozmachu nových typů rostlin. Z původně mořských řas vznikaly typy nejdříve bažinné, později již dobře přizpůsobené k životu na souši, to je mimo přímý dosah vodního prostředí.
Nejstarším prvotním suchozemským rostlinstvem byl nejprimitivnější typ rostlin tajnosnubných cévnatých, který už nemá v dnešních zástupcích této skupiny žádného příbuzného. Byla to Psilophyta, nejdůležitější skupina devonských výtrusných rostlin. Objevily se na sklonku siluru a vymizely již koncem devonu.
Ve středním devonu začala psilofytní květena pozvolna ustupovat výše organizovaným kapraďorostům, které již ve svrchním devonu vytvořily typy stromovité. Ve středním devonu se vyvinuli také zástupci rostlin přesličkovitých a plavuňovitých, takže se ráz květeny v této době již blížil rostlinstvu karbonskému.
Protože v devonu nastaly převratné změny horotvorné i klimatické, mnoho starších specializovaných živočišných skupin se novým podmínkám nedovedlo přizpůsobit a postupně vymřelo. Jiné skupiny, méně specializované, ale zato daleko schopnější se přizpůsobit, vytvořily četné nové formy.
V teplých mělkých mořích devonského údobí se dále rozvíjeli korálnatci hlavně ze skupiny korálů drsnatých a deskatých. Teprve na konci tohoto útvaru ustoupily také stromatopory, dříve tak hojně rozšířené. Koráli spolu se stromatoporami a vápenitými řasami vytvářeli během devonu četné útesy, z nichž mnohé mají dodnes velký průmyslový význam (ložiska čistých vápenců aj.).
Vrcholu svého rozvoje dosáhli v devonu ramenonožci. Byli to většinou obyvatelé mělčin a do hlubokých moří pronikli teprve v pozdějších geologických dobách. Ve značném rozvoji byly i některé skupiny měkkýšů. Byli to hlavně stočení, drobní primitivní zástupci důležité skupiny amonitů (Ammonoidea). Mlži (Lamellibranchia) i břichonožci (Gastropoda) byli v devonu zastoupeni primitivními formami, ale mlži již v tomto útvaru dosáhli prvního rozkvětu a jejich zástupci pronikali do sladkých vod.
Devonské vody, a to sladké i slané, byly velmi příhodné pro rozvoj mnoha skupin ryb. Rybovití praobratlovci (Agnatha) byli sice ještě na počátku devonu zastoupeni četnými rody, ale pozvolna vymírali, až koncem devonu zmizeli úplně. Vedle nich nastal v devonu rozvoj čelistnatých (Gnathostomata). Poprvé se objevili pancířnatí (Placodermi), kteří se sice značně podobali některým těžce obrněným bezčelistným, ale měli již zřetelné čelisti. Objevili se také předci pravých žraloků a nastal rozvoj i některých skupin ryb kostnatých (Osteichthyes), z jejichž jedné skupiny se koncem devonského útvaru vyvinuli i první obojživelníci. Skoro každá skupina ryb, která žila ve spodním devonu, však koncem tohoto útvaru vymřela. Výjimku činí pouze trnoploutví (Acanthodii), kteří žili od siluru do permu, a pak ještě některé jiné skupiny ryb kostnatých, které se koncem devonu velmi rozmohly a ve vývoji pokračovaly dále.
Skupinu pancířatých (Placodermi) objevil před více než 100 lety skotský skalník H. Miller. Našel tehdy zvláštní zkamenělinu podobnou rybě, která však měla nejen hlavu, ale zčásti i trup pokrytý kostěným pancířem a místo ploutví měla zvláštní ploutvovité výrůstky, připomínající končetiny želv.
První zástupci ryb trnoploutvých (Acanthodii) se sice objevili již v siluru, ale hlavní rozvoj nastal až v devonu. Akantodi se již mnohem více podobali současným rybám. Byli většinou drobní, kolem 10 cm dlouzí. Jejich tělo mělo protažený rybovitý tvar.
V devonských mořích žili jak předci ryb kostnatých, tak předci žraloků. Žraloci (Selachii) jsou dnes řazeni mezi paryby (Chondrichthyes), kam klademe čelistnaté obratlovce, kteří mají kostru po celý život úplně chrupavčitou, ale může být místy částečně zpevněna uhličitanem vápenatým.
Předky žraloků musíme hledat pravděpodobně mezi předky čelistnatců plakodermního typu. Žádnou skupinu plakoderm nelze však zatím považovat za skutečného předka paryb, tedy ani žraloků.
První zbytky žraloků se objevily v mořích středodevonských. Jsou to jen ojedinělé zuby popsané pod jménem Cladodus. V pozdním devonu však byly nalezeny v USA zbytky žraloka, jemuž popsané zuby náležely. Kromě nich byly nalezeny i otisky kůže, obrysy těla, stopy po měkkých tkáních (po svalech a ledvinách) i zbytky chrupavky prostoupené vápenatými solemi. Tyto zbytky byly popsány pod jménem Cladoselache. Celý jedinec dosahoval délky asi 70 cm. Je považován za představitele primitivního řádu paryb zvaného Cladoselachii. Tato primitivní skupina prastarých žraloků byla zřejmě značně blízká základní vývojové větvi všech žralokovitých.
Předci dnes běžně rozšířených ryb kostnatých (Osteichthyes) se objevili na scéně života již počátkem devonu. Brzy se však velmi rozšířili a koncem devonu již byli vedoucí skupinou obratlovců ve sladkých vodách. Protože jejich charakteristickým znakem je kostra tvořená kostní tkání, dostali vědecký název Osteichthyes - ryby kostnaté. Téměř od samého počátku jejich vývoje lze mezi nimi rozeznat tři větší skupiny, a to ryby paprskoploutvé (Actinopterygií), ryby dvojdyšné (Dipnoi) a ryby lalokoploutvé (Crossopterygií).
Ryby paprskoploutvé (Actinopterygií) jsou velmi důležité, neboť jsou nesmírně rozšířené i v současných mořích. Patří k nim totiž asi 90 % všech druhů žijících ryb. Nejcharakterističtější prvohorní skupinou těchto ryb jsou paleoniscidi (řád Palaeonisciformes). Začínají ve středním devonu a končí až ve spodní křídě.
Ryby řídkokostné (Chondrostei) zahrnují primitivní, prvohorní a triasové typy ryb i několik rodů specializovaných nebo degenerovaných, které přežily až do současnosti a patří do řádu jeseterů (Acipenseriformes).
Ryby mnohokostné (Holostei) jsou ryby charakteristické pro střední druhohory. Jejich recentní zástupci představují dožívající skupinu. Patří sem rod Lepisosteus (kostlín), žijící ve sladkých vodách Severní Ameriky a rod Amia (kaproun), obývající vody Severní a Střední Ameriky.
Ryby celokostné (Teleostei) zahrnují většinu recentních ryb, a to jak sladkovodních, tak i mořských. Objevily se koncem druhohor a od té doby jsou vládnoucí skupinou mezi rybami.
Od ryb dvojdyšných a lalokoploutvých se liší ryby paprskoploutvé četnými anatomickými znaky.
Ryby dvojdyšné (Dipnoi) představují starobylou, dnes již vymírající podtřídu zastoupenou jen několika málo druhy vyznačujícími se tím, že mohou dýchat jak žábrami, tak plícemi. Objevily se počátkem devonu a ještě během tohoto útvaru dosáhly rozkvětu.
Ryby lalokoploutvé (Crossopterygii) nejsou důležité počtem žijících druhů, ale především z hlediska vývojového. Představují totiž základní skupinu obratlovců, z níž se vyvinuly všechny další vyšší skupiny obratlovců (i člověk!).
Vývoj života dospěl tedy v devonu k důležitému mezníku - k údobí, kdy z ryb vznikl první čtvernožec, první obojživelník. První předkové obojživelníků byli skoro nerozlišitelní od ryb lalokoploutvých. Byli to poměrně dost velcí dravci, kteří žili ve vodě, kde se jim dobře dařilo. Jediný význačnější rozdíl mezi lalokoploutvými rybami a praobojživelníky je ten, že praobojživelníci měli poněkud silněji vyvinuté končetiny, lépe přizpůsobené k pohybu na souši. Když jezera nebo močály, v nichž žili, začaly vysýchat, byli předkové obojživelníků schopni vylézt po bahně vyschlých kalužin a dostat se do jiné kaluže, kde ještě voda byla.
![]()
Údobí, které následovalo po době devonské a trvalo až do konce prvohor, označují geologové souborným názvem mladší prvohory. Počítáme k nim dva útvary. První proslul mohutnými, hospodářsky nesmírně důležitými uloženinami černého neboli kamenného uhlí. Podle této významné suroviny se tento útvar nazývá karbon (carbo = latinsky uhel). Jeho trvání se odhaduje na 65 miliónů let. Druhý útvar, méně významný a kratší než karbon, se nazývá perm. Trval 55 miliónů let. Oba útvary mladších prvohor bývají v mnoha oblastech spojeny téměř neznatelnými přechody, a proto se pak označují souborně jako permokarbon.
Karbon - doba obojživelníků
Počátky karbonu charakterizují mohutné horotvorné pohyby zvané hercynské vrásnění. Během něho se od konce devonu až do počátku druhohor vyvrásnila mohutná pevninská soustava s rozsáhlými horstvy. Na mapách karbonských pevnin můžeme sledovat obrovskou severoatlantskou pevninu, která je od pevniny jižní polokoule, tehdy většinou zaledněné Gondwany, oddělena širokým středozemním mořem Tethys, které obepínalo celou Zemi. Oblast karbonského moře Tethys byla značně neklidná. Ve střední Evropě vzniklo v té době mohutné variské horstvo. Důsledkem horotvorných pohybů byla ve středních Čechách na dlouhou dobu přerušena mořská sedimentace. Horotvornou činnost doprovázela i mohutná sopečná činnost. V konečných fázích vrásnění tuhly pod povrchem vrásněných pohoří spousty magmatu a vznikaly tak mohutné masívy žul i jiných hornin. V té době vznikly žulové masívy, jako např. středočeský, krkonošsko-jizerský aj. Ve svrchním karbonu začaly v několika pánvích i výlevy porfyrů a melafyrů, které však vrcholily až v permu.
V karbonu tedy vzniklo na prostorách dříve pokrytých devonskými moři a později spodnokarbonskými moři a močály mohutné horstvo variské. Na jeho okrajích v četných lagunách a deltách řek bujelo rostlinstvo a na mnoha místech se hromadilo obrovské množství rostlinné hmoty, z níž pozvolnými chemickými pochody, jimž říkáme uhelnatění vznikala rozsáhlá ložiska černého uhlí. Mořská hladina místy kolísala a mořská voda znovu v určitých dobách zaplavovala uhelné močály. Když moře ustoupilo, vznikaly nové pralesy - a pochod se opakoval. Uvnitř variského pohoří vznikaly místy ve vnitrozemských pánvích rozsáhlé močály, které však moře nezaplavovalo a v nich se ukládalo výborné černé uhlí (u nás např. kolem Plzně a jinde).
Konec karbonu je místy dost nejasný a v praxi se vymezuje hlavně podle zkamenělin. Charakteristickou horninou karbonu je kamenné uhlí. Protože příznivé podmínky pro rozvoj rostlinstva nenastávaly všude stejně, vznikaly uhelné sloje v různých částech světa v různých dobách. Ohromný rozvoj rostlinstva svědčí o tom, že karbonské podnebí v oblasti Evropy bylo většinou vlhké a teplé. Rovněž tehdejší středozemní moře Tethys bylo podle bujného růstu vápenitých řas a výskytu četných druhů bezobratlých velmi teplé.
Suchozemské rostlinstvo dosáhlo již v druhé polovině devonu určitého rozmachu. Bylo vlhko, teplo a ve vzduchu byl dostatek oxidu uhličitého. To pomáhalo nejen vývoji nových rostlinných druhů, ale podporovalo i neobyčejně bujný a často stromovitý růst. Vznikaly obrovské pralesy. Většinou se však omezovaly na močálovité a jezerní pánve, protože rostlinstvo karbonské doby patřilo převážně do skupiny rostlin tajnosnubných cévnatých a nedosáhlo ještě toho stupně dokonalosti, aby mohlo rovnoměrně osídlit i suchá místa. Mezi starobylou tajnosnubnou květenou, pro niž jsou charakteristické především stromovité plavuně a přesličky, vyrůstali již také první průkopníci rostlinstva výše organizovaného, pokročilejšího, které v té době dobylo i sušších míst, přizpůsobilo se jim a natrvalo je osídlilo; byly to přední hlídky rostlin nahosemenných.
Ze stromovitých plavuní jsou nejznámější dva typy, Lepidodendron a Sigillaria. Kmeny rodu Lepidodendron dosahovaly výšky až 30 m. Koruny byly bohatě rozvětvené. Při zemi dosahoval kmen až 2 m šířky a přecházel ve vidličnatě se větvící systém výrůstků podobných kořenům, které upevňovaly strom v bahnité půdě nebo i v pobřežním bahně jezer. Listy byly v příčném průřezu široce kosočtverečné, několik centimetrů až 1 m dlouhé a pokrývaly jen mladé větve. Kmen a staré větve byly bezlisté. Na koncích větví byly výtrusnicové šištice. Povrch kmenu byl pokryt listovými polštářky s listovými jizvami, které jsou pro rod Lepidodendron charakteristické.
Jiným typem stromovitých plavuní byl rod Sigillaria. Přes 20 m vysoké kmeny těchto plavuní byly zakončeny chvostem úzkých dlouhých listů. Kmen se na konci dělil ve dvě nebo více větví, z nichž každá byla opět zakončena chvostem dlouhých listů. Výtrusnicové šištice, někdy až 30 cm dlouhé a 7 cm široké, byly dlouze stopkaté a vyrůstaly jednotlivě či ve skupinách z kmene nebo z bezlistého spodku vidličnaté koruny. Také tyto kmeny nesly na svém povrchu jizvy po odpadlých listech. Plavuně rodu Sigillario byly pro karbon velmi charakteristické.
Ke stromovitým plavuním se družily i stromovité přesličky. Byly to rovněž vysoké stromy s kmenem i větvemi ostře článkovanými. Přesličky měly rády vlhko, a proto vyrůstaly obvykle z vod jezer, bažin a močálů. Jejich kmeny byly stejně jako stonky dnešních přesliček duté. Listy byly úzké, čárkovité a vyrůstaly na větvích v přeslenech. Do přeslenů byly na kmenech sestaveny i větve.
Nejkrásnější a také nejpestřejší složkou porostů kamenouhelných pralesů byly bezesporu kapradiny. Jejich zuhelnatělé listové vějíře, složené z lístečků nejrozmanitějších tvarů a velikostí, nalézáme velmi často v různých uhelných lupcích a břidlicích.
V kamenouhelných pralesích zvlášt vynikaly četné stromovité kapradiny. Mohutné vějíře jejich listů vyrůstaly buď jen ze dvou stran kmene, nebo z celého jeho obvodu. Kmeny těchto kapradin byly obaleny hustě propletenými kořínky a vyrůstaly do výše 10-15 m. Teprve v této výšce se rozkládaly honosné živě zelené koruny. Stromovité kapradiny vyrůstaly spíše na místech sušších než vlhkých.
V kamenouhelných pralesích rostly i jiné rostliny blízké nahosemenným, a to tzv. kordaity. Byly to stromy vysoké až 30 m, se štíhlým kmenem, nepravidelně rozvětveným v korunu. Měly široké pentlicovité listy se souběžnou žilnatinou, které vyrůstaly na větvích ve šroubovici. Po opadání listů zůstávaly na větvích eliptické jizvy. Květy tvořily řídké různopohlavné šištice. Kordaity pravděpodobně patřily ve své době mezi největší stromy vůbec.
Jinými velikány kamenouhelných pralesů byly i nejstarší nahosemenné rostliny lebachie (Lebachiaceae). Patří k nim první typické jehličiny. Vyskytují se sice od svrchního karbonu, ale největšího vývoje dosáhly až ve spodním permu. Vyhledávaly spíše suchá místa, a proto nalézáme jejich zbytky mezi vlhkomilným rostlinstvem karbonských močálů jen vzácně.
V karbonských mořích se velmi rozšířili prvoci ze skupiny dírkovců (Foraminifera). Některé jejich rody dosahovaly nápadné velikosti a vyskytovaly se v takovém množství, že se staly důležitými horninotvornými činiteli. Pro dělení karbonských vrstev mají velký význam hlavně koráli a ramenonožci.
Vrcholu svého rozvoje dosáhly lilijice. Z měkkýšů zaslouží pozornost hlavonožci, zvláště goniatiti, kteří měli mnoho druhů důležitých pro určování a srovnávání mořských vrstev. Jiná skupina měkkýšů, břichonožci, pronikla v této době i do sladkých vod. Trilobiti již zvolna vymírali.
Karbon se vyznačoval pozoruhodným rozvojem suchozemských členovců, hlavně různých čeledí hmyzu, stonožek, štírů, pavouků aj. Nejhojnější byl hmyz, který je i dnes počtem druhů a jedinců jednou z nejrozšířenějších tříd živočichů.
Ve spodním karbonu není po zkamenělém hmyzu téměř ani stopy, ale ve svrchním karbonu byl celý tehdejší kontinent "náhle" hmyzem přímo zaplaven.
Nejzajímavější z těchto vymřelých skupin byli pramřížonerví (Palaeodictyoptera), jediný hmyz se šesti křídly.
Různorodou a ohromnou třídu hmyzu dělíme dnes na dvě velké skupiny: bezkřídlé (Apterygota) a křídlaté (Pterygota).
Ve vývoji hmyzích křídel můžeme předpokládat čtyři stadia. Stádium skákání, plachtění, veslování a stádium skutečného letu.
U některých skupin (Palaeoptera) se mohla křídla pohybovat pouze nahoru a dolů. Schopnost ohýbat křídla a v době odpočinku je skládat na zadečku znamenala krok kupředu. Umožnila hmyzu lepší skrývání i lepší možnost snášet vajíčka do chráněných míst.
Vedle hmyzu žilo v karbonských pralesích mnoho dalších skupin členovců. Pod tlejícími zbytky rostlinstva se skrývalo množství pestře zbarvených velkých stonožek (Chilopoda) a pod kameny a kmeny padlých a hnijících stromovitých plavuní i jiných stromů se ukrývali různí jedovatí štíři (Scorpionida). V nejrůznějších úkrytech čekaly na svou kořist četné druhy pavoukovců (Arachnida).
Podivuhodným a zároveň největším členovcem karbonských pralesů byla obrovská Arthropleura. Podobala se dlouhé stonožce. Její tělo bylo úzké, protáhlé, článkované a dosahovalo délky až 150 cm. Tito vymřelí členovci žili ve vlhkých lesích a byli patrně všežravci.
Velmi bohatý byl život ve vodách karbonských jezer a močálů. V bahnitém dně žili různí červi, v čistých vodách mlži, po stoncích vodních rostlin lezli vzácně také někteří břichonožci. Ve vodách byli hojní nejrůznější drobní korýšci a vzácněji se na některých místech vyskytovali i hrotnatci (Merostomata), pradávní předkové ostrorepů.
Rybí fauna sladkých vod v karbonu se podstatně lišila od fauny devonské. Velmi rozšíření byli drobní akantodi, dosahující délky maximálně 30 cm. Na území ČSSR se hojně vyskytoval rod Acanthodes. V karbonských jezerech a řekách žili také velcí pražraloci z řádu Xenacanthida. Měli chrupavčitou kostru a těsně za hlavou před hřbetní ploutví nápadný, mohutný trn. Ocasní ploutev byla nesouměrná, tlama byla vyzbrojena četnými špičatými zuby. Dosahoval délky až 70 cm. Poměrně hojné byly v karbonských vodách také ryby kostnaté, a to dvojdyšné i paprskoploutvé.
Velký rozmach bezobratlých živočichů, a zvláště členovců v karbonských pralesích silně podpořil také rozvoj primitivních obojživelníků - krytolebců neboli stegocefalů. Krytolebci vznikli z lalokoploutvých ryb a na karbonských pevninách, kde téměř neměli nepřátel, se nesmírně rozmnožili.
Krytolebci dosahovali různých velikostí. Někteří měřili jen několik málo centimetrů, jiní dosáhli velikosti menších aligátorů. Někteří se podobali ještěrkám, jiní mlokům nebo hadům, a někteří dokonce i krokodýlům. Jejich tělo bylo pokryto pancířem z jemných šupin kruhovitého, oválného nebo protaženého tvaru. Hlava byla plochá, široce trojbokého tvaru nebo tvaru gotického oblouku. Tlama byla vyzbrojena ostrými špičatými zuby prozrazujícími, že krytolebci byli dravci. Od dnešních obojživelníků se lišili stavbou lebky, kterou tvořil mnohem větší počet kostí, jež lebku úplně pokrývaly. Odtud také pochází jejich český název: krytolebci. Mezi týlními kostmi měli krytolebci zřetelné třetí oko, které sloužilo jako světločivý orgán.
Krytolebci obývali vlhké břehy jezer a klidně tekoucích řek, ale hlavně močály a tůně karbonských pralesů. Množili se vajíčky, která snášeli do vody a z nichž se líhly larvy dýchající vnějšími žábrami.
Jak jsme se již zmínili, koncem karbonu, a zvláště pak na počátku permu se začalo vlhké a teplé podnebí měnit, období sucha se prodlužovala, až posléze v permu suché podnebí úplně převládlo. Již koncem karbonu ubylo vlhkých pralesů a zvětšily se suché pustiny. Tyto hluboké změny měnícího se prostředí měly samozřejmě pronikavý vliv na vývoj živočišstva, především obratlovců. Krytolebci se museli stáhnout do oblastí, kde se ještě udržovala voda, ale i tam je postupující sucho hubilo. Mezi rody, které ještě nebyly příliš specializované, začaly se objevovat populace, které se přizpůsobovaly životu v sušších oblastech. Na povrchu jejich těl se objevila zrohovatělá pokožka, tvořící pevné krunýře, které dobře ochraňovaly tělo před rychlým vypařováním vody. Tyto formy přestaly snášet vajíčka do vody a zahrabávaly je do hromad tlejícího rostlinstva nebo přímo do písku. Vodu, nezbytnou k vývoji zárodků, nahrazovala tekutina v tzv. amnionové dutině vajíček. Toto zařízení zůstalo až dodnes u plazů, ptáků a savců, tedy u obratlovců, které zahrnujeme do skupiny Amniota. Vedle těchto změn se v základě změnila i stavba kostry. Tak vznikla nová, velmi důležitá skupina zvířat - první plazi. Z hlediska historického vývoje organismů je vznik plazů a jejich úplný přechod na souš významným pokrokem.
Ačkoliv se první plazi objevili už ve svrchním karbonu, dosáhli teprve ve spodním permu většího rozvoje a prvního rozkvětu. V této době už je možné rozeznat mezi nimi několik základních větví. Fosilní zbytky nejstarších karbonských plazů jsou však velmi vzácné. Poněkud hojnější jsou zbytky z vývojového hlediska důležitých savcovitých plazů - pelykosaurů, jejichž zástupce Edaphosaurus s mohutně protaženými obratlovými trny byl nalezen i na území ČSSR.
Perm
Posledním útvarem prvohor je perm. Byl kratší než karbon, trval asi 55 miliónů let. Horotvorné pochody hercynské doznívaly. Jiné životní podmínky byly v první polovině tohoto útvaru a jiné zase v jeho druhé polovině. Měnilo se podnebí i povaha prostředí. Starobylé typy květeny a zvířeny vystřídaly typy mladší, modernější, novější. Život rychle pokračoval ve svém vývoji ve vodě, na souši i ve vzduchu. Tímto útvarem končí prvohory a zároveň se v něm rodí druhohory.
Zvětšení souší a stoupání průměrné nadmořské výšky byly hlavní příčinou změny podnebí. Na severní polokouli, kde se šířil daleko k severu pás teplého podnebí, utvořila se rozsáhlá, poměrně suchá pustinná oblast. Vlhčí mírné podnebí se přesunovalo dále k východu do oblasti uralské, kde nastaly teprve teď příhodné podmínky pro vznik uhelných slojí. Velké části jižní prapevniny Gondwany byly zaledněny.
Ve spodním permu ještě převládaly rostliny výtrusné a kapraďosemenné jako v karbonu, ale ve svrchním permu už nabyly převahy rostliny nahosemenné. Mezi nimi se již objevily typy, které potom rostly hojně v druhohorách. Ve složení květeny svrchního a spodního permu je nápadný rozdíl. Na rozhraní těchto dvou permských údobí končí totiž starší období vývoje rostlin (paleofytikum) a začíná období střední (mezofytikum), jež se vyznačuje převahou nahosemenných.
Ve spodním permu mizely z dosavadní společnosti stromovitých rostlin výtrusných plavuně rodu Lepidodendron; sigillárie se udržovaly jen v několika typech. Z kapradin a rostlin kapraďosemenných se objevily některé nové druhy. Zkřemenělé kmeny kapradin s velmi dobře zachovanými tkáněmi známe pod jménem psaronie. Ve spodním permu doznívaly i kordaity. Začaly se objevovat některé typy jinanovitých a cykasovitých rostlin. Nejvíce se však v tomto sušším období rozšířily rostliny jehličnaté. Na jižní polokouli se rozšířila gondwanská flóra glossopterisová.
Zatímco rostlinstvo dosáhlo vyššího stupně vývoje již ve svrchním permu, vývoj živočišstva byl v tomto směru poněkud opožděn; nového vývojového stupně dosáhli živočichové až koncem tohoto útvaru.
Vůči karbonu pozorujeme v permu jen málo změn. V mořské fauně se dále vyskytují dírkovci, pozvolna mizí čtyřčetní koráli a svůj poslední velký význam mají ještě ramenonožci. Poměrně hojní jsou mlži; jejich četné rody mají již blízké vztahy k fauně druhohorní. Vedle posledních goniatitů se vyskytují již i první praví amoniti. Velmi hojní byli ostnokožci, hlavně poupěnci a lilijice. Z obratlovců se rozvíjejí hlavně plazi, kdežto ostatní obratlovci, např. některé skupiny ryb a obojživelníků jsou na ústupu, popřípadě vymírají. Permské ryby se od ryb karbonských mnoho nelišily. Ve sladkých vodách se stále hojně vyskytoval rod Xenacanthus a drobný Acanthodes. Velkého rozvoje však dosáhl rod Palaeoniscus, který je pro perm typický. Předkové tohoto rodu se objevili poprvé ve sladkých vodách středního devonu.
V permu žilo v různých částech světa mnoho čeledí starobylých obojživelníků (krytolebců). Většina jich však koncem permu vymřela. Do triasu přežily jen nečetné rody. Některé druhy dosáhly v té době značně velkých rozměrů, až velikosti aligátora.
Vývojově nesmírně důležitou skupinou, která se v permu již značně rozšířila, jsou plazi. Vyvinuli se z pokročilých typů obojživelníků během karbonu a poměrně rychle se stali vládnoucí skupinou v druhohorách. Postupně z nich vznikaly nejrůznější a nejpodivnější typy často obludných rozměrů, avšak koncem druhohor začaly rychle vymírat. Jejich zánik, i když zpomaleně, pokračuje dodnes. V době největšího rozmachu, v druhohorách, však z nich vznikli savci a ptáci, kteří se pak během dalších geologických dob značně rozmohli.
První zástupci plazů z konce karbonu jsou velmi vzácní a obvykle špatně zachovaní. Teprve v permu, kdy změnou klimatu vznikly rozsáhlé vyprahlé oblasti a kdy obojživelníci ustupovali z vrcholu své slávy, nastala doba, v níž se plazům začalo výborně dařit.
V poměrně krátké době vzniklo mnoho nových forem, z nichž většina patří ke dvěma význačným řádům, Captorhinomorpha a Pelycosauria.
Captorhinomorpha byli většinou velcí, těžkopádní plazi s četnými znaky upomínajícími ještě na krytolebce. Byla to nejprimitivnější skupina plazů. Lebku měli úplně uzavřenou kostmi, tělo bylo těžkopádné, nemotorné. Existovaly již formy býložravé a masožravé. V permu byli značně rozšířeni a vytvořili mnoho různých typů. Přizpůsobením k příslušnému prostředí z nich brzy vznikla řada jiných skupin.
Vrchol rozvoje řádu Captorhinomorpha probíhal v permu. V triasu jeho poslední příslušníci vymřeli.
Jiná pozoruhodná skupina plazů, která se objevila ve svrchním permu jižní Afriky, byli předkové ještěrek a pozdějších hadů, řád Eosuchia. Patří k němu nevelcí, velmi pohybliví draví ještěři s lehce stavěnou kostrou. Měli dva spánkové otvory, dvouduté obratle a velmi primitivně stavěné končetiny. Skupina Eosuchia vznikla pravděpodobně z primitivních kaptorhinomorfních plazů a v triasu se z ní vyvinul řád Rhynchocephalia, jehož primitivní zástupce, tříoká haterie (Sphenodon), žije dodnes na Novém Zélandě; je to skutečná žijící fosilie. Ostatní zástupci tohoto řádu vymřeli během druhohor.
Druhá velká skupina primitivních plazů z konce karbonu a permu patří mezi "plazy savcovité" - (Synapsida). Jsou to pelykosauři (Pelycosauria), od nichž lze sledovat vývojovou linii již přímo k počátkům savců a člověku. Savci se ujali vlády nad živočišným světem poměrně pozdě.
Druhou větev představuje býložravý rod Edaphosaurus. Měl tělo zavalitější a kratší než Varanosaurus a rovněž lebku měl poměrně malou. Byl to přes 3 m dlouhý býložravec. Velmi charakteristické pro tento rod byly protažené trnové výběžky obratlů, vybíhající vysoko nad hřbet, s nápadnými příčnými výrůstky.
Třetí vývojovou větev charakterizuje přes 3 m dlouhý masožravý plaz rodu Dimetrodon. který žil ve spodním permu Severní Ameriky. Podobal se rodu Edaphosaurus, ale neměl příčné výrůstky na silně protažených trnových výběžcích obratlů. Měl ostré zuby s nápadnými trháky a byl již úplně přizpůsoben životu na souši.
Štítky
Nebyly nalezeny žádné štítky.